
F因子(F factor)は細菌の環状染色体との間で交叉を行うことで、染色体中に組み込まれます。
F因子には、挿入配列(insertion sequence)と呼ばれるそれぞれ約1000塩基の長さのDNA領域が数個あります。この挿入配列は細菌の染色体の似た場所と対合し、交叉を起こすことができます。細菌のゲノムにはF因子が入り込むことができます。
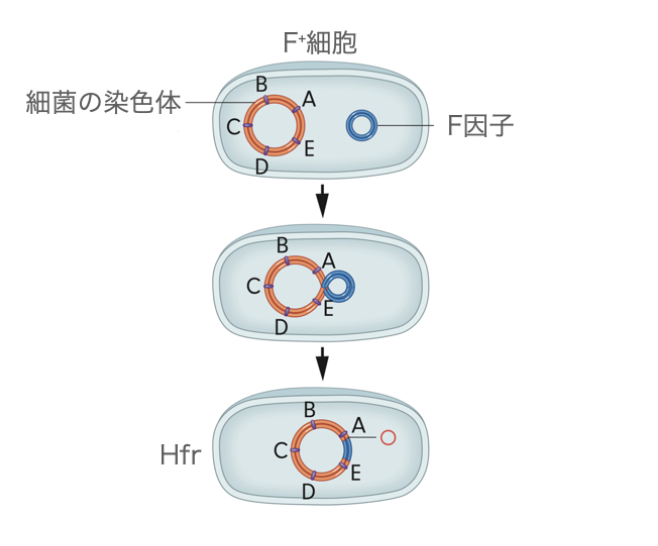
F因子が染色体に組み込まれた細菌は高頻度組換え(high frequency of recombination)と呼ばれ、一般に略してHfrと呼ばれます。Hfr細胞になった細菌は、その後の細胞分裂では組み込まれたF因子も含めて自己複製します。
そして、F⁻細胞へF因子の移行が行われるときと同様に、遺伝子の以降を行います。この場合、細菌の染色体の一部が移行します。また、組み込まれたF因子もF⁺と同様に接合の際に必要な線毛の形成を行います。
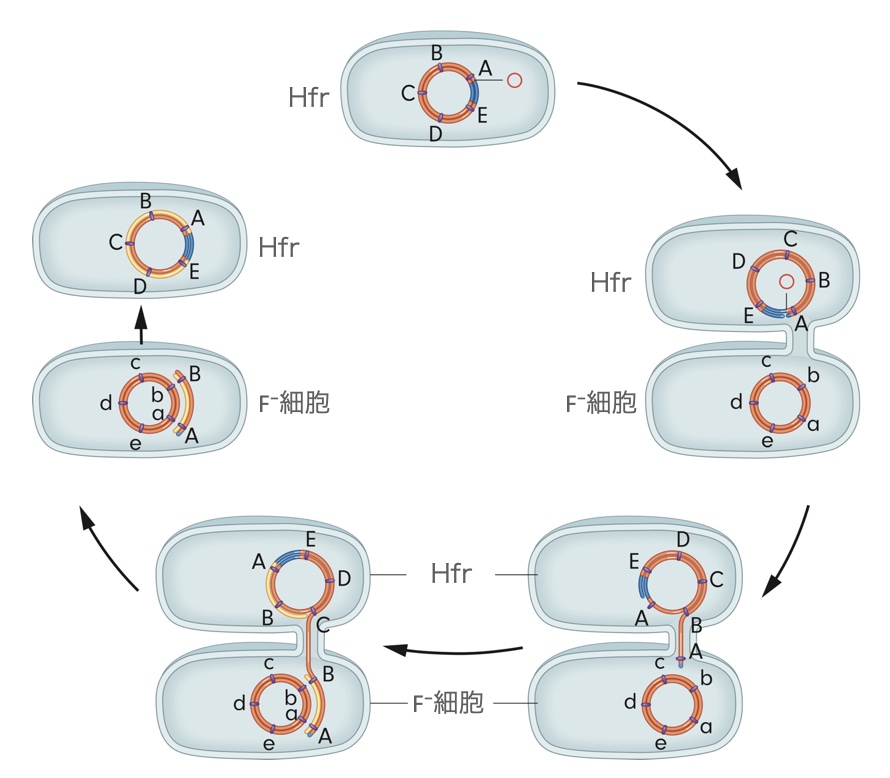
Hfrの染色体がF⁻に移行される仕組み
Hfrの染色体には組み込まれたF因子の部分(上図では青の部分)があります。そして、元来、F因子のDNAであった領域に切れ目となる部分があります。そして、そこからF⁻へ伸びた接合管から、Hfrの染色体が移行されます。
切れ目のすぐ隣にある遺伝子をAとし、そこから順番にB・C・D・Eと遺伝子に名前をつけていきます。すると、常に、A⇒B⇒C⇒D⇒E⇒Fの順番で、F⁻因子へと移行が起こることになります。また、f因子が組み込まれた方向によってはE⇒D⇒C⇒B⇒A⇒Fの順番のこともあります。もし、BとCの間にF因子が組み込まれた場合、C⇒D⇒E⇒A⇒B⇒Fの順番になります。
DNAの単鎖だけが接合管を通ります。そして、F⁻の細胞の中で相補鎖が合成されます。このときは、Hfr由来の染色体は棒状の二重鎖です。この棒状二重鎖の長さは交配が行われた時間によって変わります。上図ではHfrのAーB間だけがF⁻細胞に移行された場合です。
そして、この棒状二重鎖は、F⁻染色体と交叉します。移行したHfrの遺伝子のどれが、F⁻細胞の染色体に恒久的に組み込まれるかは、移行したDNAのどの部分で交叉が起きるかによって異なります。そして、この際の交叉で、F⁻の染色体に組み込まれることになるのは、二重や四重といった偶数回の交叉が起こった場合です。奇数回の交叉は、F⁻染色体の環状構造を破壊してしまいます。
一方、Hfr細胞の側では、残った単鎖が鋳型となって相補鎖が形成され、環状二重鎖になります。移行は単鎖によって行われます。
F’因子 – 細菌のDNAの一部をもったF因子
交叉によって染色体に組み込まれることができるF因子ですが、これと逆の過程が起こることで、HfrはF⁺因子に逆戻りすることができます。そして、このような交叉が起こったときには、F因子は完全に元々組み込まれた部分だけで出てくるわけではなく、細菌のDNAのいくつかをもって出てきます。このような細菌の遺伝子をもったF因子はF'(Fプライム)と呼ばれます。
F’が染色体に組み込まれてHfrを形成する場合は、F’は常に同じ場所に組み込まれます。F’の中の細菌DNAとの相同性のためだと考えられています。
F’系統を用いることで、局所的には二倍体である状態を作ることができます。二倍体の場合、遺伝子の優越関係を研究することができますが、通常、大腸菌は半数体です。そのため半数体の大腸菌では優劣関係の研究ができません。F’因子の中で最もよく知られているものは、ガラクトースに関係する大腸菌の遺伝子です。
